
Alexander J. Varshavsky

PDF of Varshavsky's complete CV, 2023.
PDF of 2007 article about using DNA deletions for targeting cancer cells (Varshavsky (2007) Targeting the absence: homozygous DNA deletions as immutable signposts for cancer therapy. PNAS 104, 14935-14940).
PDF of the follow-up 2023 article on the same subject (Varshavsky, A., Lewis, K. and Chen, S. J. (2023) Deletions of DNA in cancer and their possible use for therapy. BioEssays 2300051).
PDF of 2008 "Reflections" article (Varshavsky, A. (2008) Discovery of cellular regulation by protein degradation. J. Biol. Chem. 283, 34469-34489).
PDF of 2014 article about the early history of the ubiquitin field (Varshavsky, A. (2014) Discovery of the biology of the ubiquitin system. J. Am. Med. Assoc. (JAMA) 311, 1969-1970 ).
PDF of 2019 "Perspective" article (Varshavsky, A. (2019) N-degron and C-degron pathways of protein degradation". PNAS 116, 358-366 ).
PDF of 2021 interview, about life and work, to David Zierler, Caltech Heritage Project.
Academic Appointments and Education:
1970: B.S. in Chemistry, Moscow University, Moscow, Russia.
1973: Ph.D. in Biochemistry, Institute of Molecular Biology, Moscow.
1973-1976: Research Fellow, Institute of Molecular Biology, Moscow.
1977-1992: Assistant, Associate, Full Professor, Dept. of Biology, MIT, Cambridge, MA.
1992-2017: Smits Professor of Cell Biology, Division of Biology, California Institute of Technology (Caltech), Pasadena, CA.
2017-present: Thomas Hunt Morgan Professor of Biology, Division of Biology and Biological Engineering, Caltech, Pasadena, CA.
Honorary Memberships:
Fellow, American Academy of Arts and Sciences, 1987.
Member, U.S. National Academy of Sciences, 1995.
Fellow, American Academy of Microbiology, 2000.
Foreign Associate, European Molecular Biology Organization, 2001.
Member, American Philosophical Society, 2001.
Fellow, American Association for Advancement of Science, 2002.
Foreign Member, European Academy of Sciences (Academia Europaea), 2005.
Scientific Awards:
Merit Award, National Institutes of Health, 1998.
Novartis-Drew Award in Biomedical Science, Novartis, Inc. and Drew University, 1998.
Gairdner International Award, Gairdner Foundation, Canada, 1999 (with A. Hershko).
Sloan Prize, General Motors Cancer Research Foundation, 2000 (with A. Hershko).
Lasker Award in Basic Medical Research, Albert and Mary Lasker Foundation, 2000 (with A. Hershko and A. Ciechanover).
Shubitz Prize in Cancer Research, University of Chicago, 2000.
Hoppe-Seyler Award, Society for Biochemistry and Molecular Biology, Germany, 2000.
Pasarow Award in Cancer Research, Pasarow Foundation, 2001.
Max Planck Award, Germany, 2001.
Merck Award, American Society for Biochemistry and Molecular Biology, 2001 (with A. Hershko).
Wolf Prize in Medicine, Wolf Foundation, Israel, 2001 (with A. Hershko).
Massry Prize, Massry Foundation, 2001 (with A. Hershko).
Horwitz Prize, Columbia University, 2001 (with A. Hershko).
Wilson Medal, American Society for Cell Biology, 2002 (with A. Hershko).
Stein and Moore Award, Protein Society, 2005 (with A. Hershko).
March of Dimes Prize in Developmental Biology, March of Dimes Foundation, 2006.
Griffuel Prize in Cancer Research, Association for Cancer Research, France, 2006.
Gagna and Van Heck Prize, National Foundation for Scientific Research, Belgium, 2006.
Weinstein Distinguished Award, American Association for Cancer Research, 2007.
Schleiden Medal, German Academy of Sciences (Leopoldina), 2007.
Gotham Prize in Cancer Research, Gotham Foundation, 2008.
Vilcek Prize in Biomedical Research, Vilcek Foundation, 2010.
BBVA Foundation Award in Biomedicine, BBVA Foundation, Spain, 2011.
Otto Warburg Prize, Society for Biochemistry and Molecular Biology, Germany, 2012.
King Faisal International Prize in Science, King Faisal Foundation, Saudi Arabia, 2012.
Breakthrough Prize in Life Sciences, Breakthrough Foundation, 2014.
Albany Prize in Medicine and Biomedical Research, Albany Medical Center, Albany, NY, 2014.
Grande Médaille, French Academy of Sciences, 2016.
Wieland Prize, Boehringer Ingelheim Foundation, Germany, 2017.
IUBMB Medal, International Union of Biochemistry and Molecular Biology, 2019.
Debrecen Award in Molecular Medicine, University of Debrecen, Hungary, 2022.
Hope Award in Basic Science, Hope Funds for Cancer Research, Newport, RI, 2023.
Hogg Award, M. D. Anderson Cancer Center and University of Texas, Houston, TX, 2023.


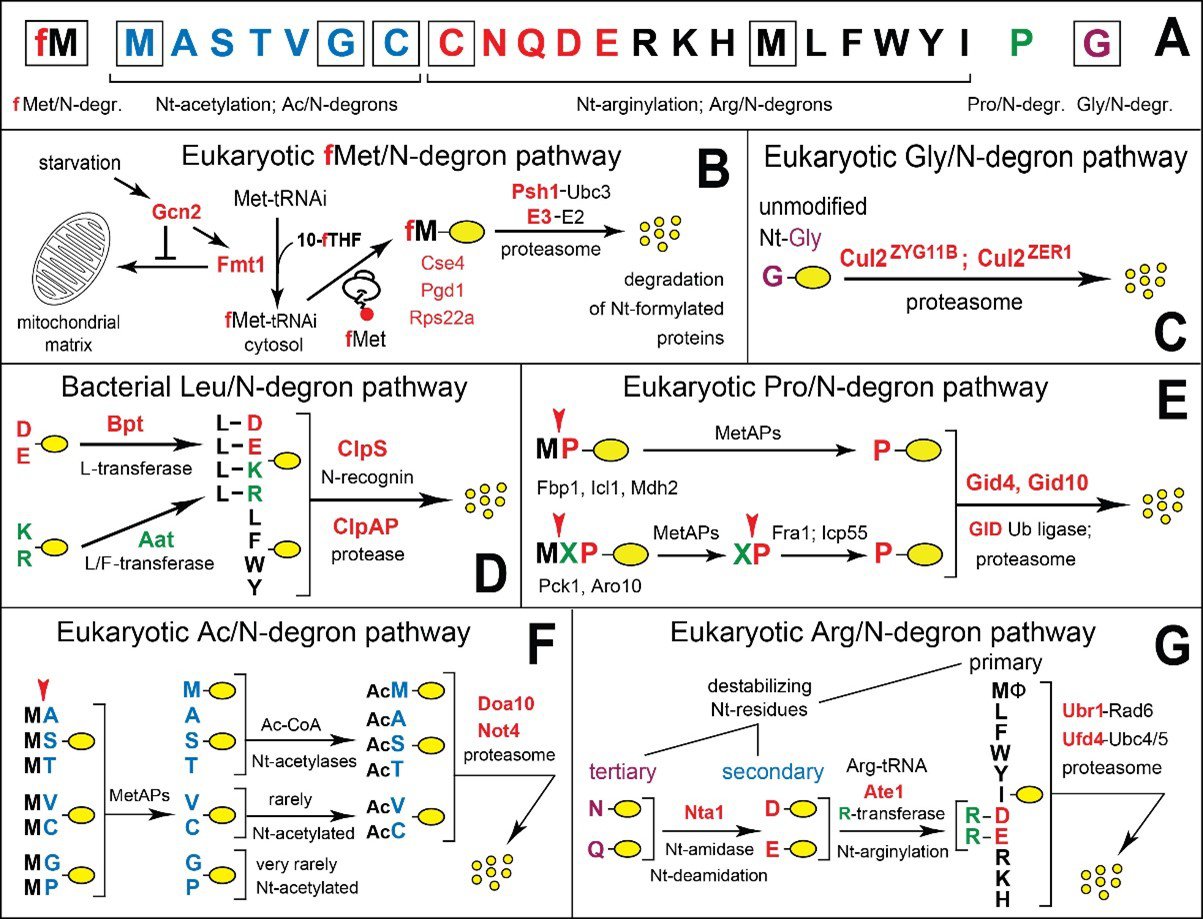
Figure 1. N-degron pathways. (A) 20 amino acids of the genetic code (as N-terminal (Nt) residues) are indicated by single-letter abbreviations. A yellow oval denotes the rest of a protein substrate. The twenty amino acids are arranged to delineate specific N-degrons. Nt-Met is cited thrice, since it can be recognized by the Ac/N-degron pathway (as Nt-acetylated Ac-Met), by the Arg/N-degron pathway (as unacetylated Nt-Met), and by the fMet/N-degron pathway (as Nt-formylated fMet). Nt-Cys is cited twice, since it can be recognized by the Ac/N-degron pathway (as Nt-acetylated Cys) and by the Arg/N-degron pathway (as the arginylatable Nt-Cys sulfinate or Nt-Cys-sulfonate). (B) The eukaryotic (S. cerevisiae) fMet/N-degron pathway. 10-fTHF, 10‑formyltetrahydrofolate. (C) The eukaryotic (mammals) Gly/N-degron pathway. (D) The bacterial (V. vulnificus) Leu/N-end rule pathway. (E) The eukaryotic (S. cerevisiae) Pro/N‑degron pathway. (F) The eukaryotic (S. cerevisiae) Ac/N-degron pathway. (G) The eukaryotic (S. cerevisiae) Arg/N-degron pathway. See papers cited below for references and other details.


{kind=link}
{kind=link}
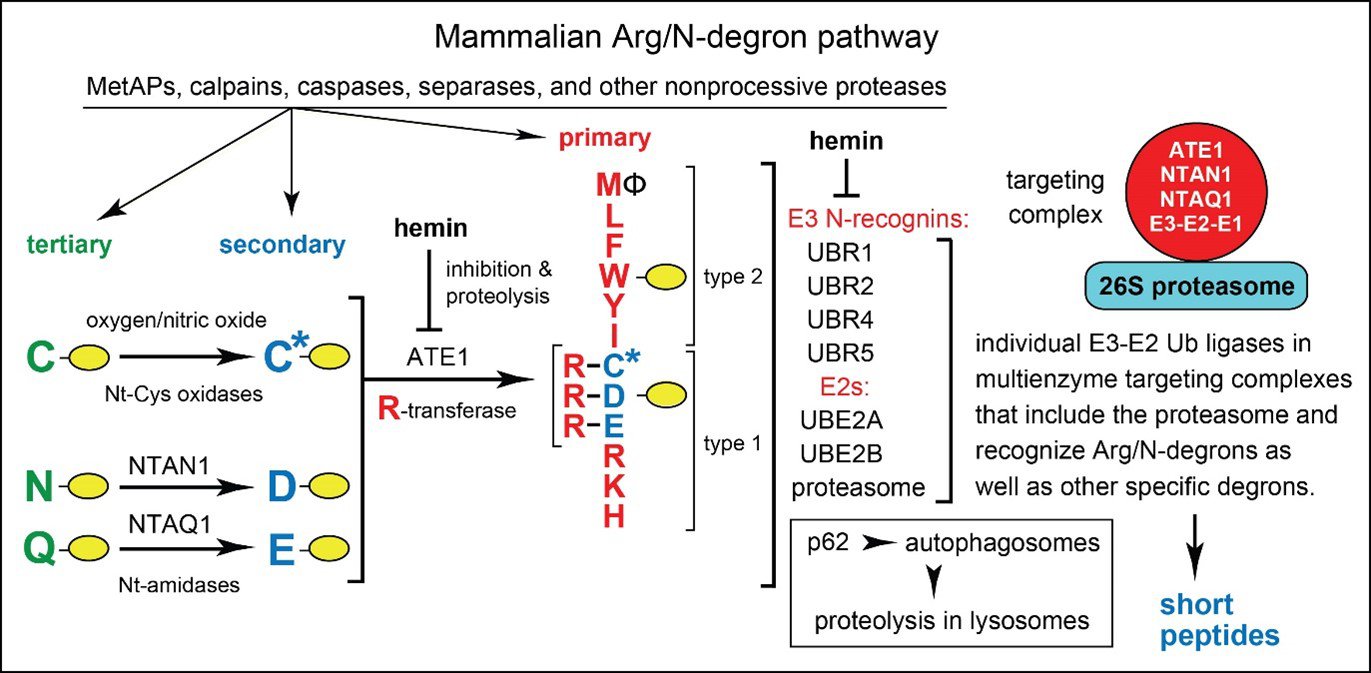
Figure 2. The mammalian Arg/N-degron pathway. This pathway targets proteins either for the proteasome-mediated degradation (via UBR1, UBR2, UBR4, and UBR5 E3 N-recognins) or lysosome-mediated degradation (via p62 N-recognin). The same designations as in Fig. 1. The cited E3 ubiquitin (Ub) ligases of this pathway can recognize not only the indicated (destabilizing) Nt-residues but also specific non‑N‑terminal degrons in proteins that lack Arg/N‑degrons. C* denotes oxidized Nt-Cys, either Nt-Cys‑sulfinate or Nt-Cys-sulfonate. The ATE1 Arg-tRNA-protein transferase (R-transferase) conjugates Arg to Nt-Asp, Nt-Glu or (oxidized) Nt-Cys. Hemin (Fe3+-heme) inhibits the enzymatic activity of R‑transferase and accelerates its degradation in vivo. Hemin also binds to UBR1/UBR2 and modulates specific aspects of their activity. Type 1 and type 2 refer, respectively, to two sets of primary destabilizing Nt-residues, basic (Arg, Lys, His) and bulky hydrophobic (Leu, Phe, Trp, Tyr, Ile, and also Met, if the latter is followed by a bulky hydrophobic residue (Ф)). Five enzymes of the mammalian Arg/N-degron pathway form a targeting complex, which apparently includes the 26S proteasome as well. See papers cited below for references and other details.
Our Studies of the Ubiquitin System
We are interested in just about everything. But the brevity of life being what it is compels selectivity. Hence our main subject – the ubiquitin system – chosen partly through a strong preference and partly by accident.
The field of ubiquitin (Ub) and regulated protein degradation was created in the 1980s, largely through complementary discoveries by the laboratory of A. Hershko (Technion, Haifa, Israel) and by my laboratory, then at MIT. These discoveries, during the 1980s, revealed three sets of previously unknown facts:
1) That the previously known ATP-dependent degradation of proteins in extracts from rabbit reticulocytes involves a new protein modification, the covalent conjugation of Ub, a 76-residue protein, to other proteins in these extracts, including proteins added to an extract. Ub-protein conjugation was shown, by the Hershko laboratory, to be mediated by specific enzymes, termed E1, E2 and E3. These studies were carried out using in vitro (cell-free) systems and purified or partially purified E1-E3 enzymes.
2) That the selectivity of ubiquitin conjugation to other proteins is determined by the presence of specific degradation signals (degrons) in cellular proteins, including N-terminal (Nt) degrons, The latter were the first degradation signals to be discovered (in 1986) and were called N-degrons. The main determinant of an N-degron in a protein is its destabilizing Nt-residue. Ub-dependent proteolytic systems that target proteins bearing specific N-degrons are called N-degron pathways. (Until 2019, these systems were called "N-end rule pathways".) The term "degron" was proposed by us in 1991 (Varshavsky, A. (1991) Naming a targeting signal. Cell 64, 13-15) and became a standard term in the field. See Figures 1 and 2, and also 2019‑2022 papers cited below, in the list of selected papers by the lab.
3) That ubiquitylation of proteins has strikingly broad biological functions, largely through control, via processive proteolysis, of the in vivo levels of most cellular proteins, and also through pathways that involve Ub but do not involve proteolysis. Specifically, during the 1980s my lab has discovered, using genetic and biochemical methods, that Ub conjugation is essential for the bulk of protein degradation in vivo (in living cells). In addition to identifying the first degrons in short-lived proteins (item 2 above), we also discovered the first specific functions of ubiquitylation: in DNA repair (1987); in the cell cycle (1988); in stress responses (1987); in protein synthesis (1989); and in transcriptional regulation (1990). We also identified the first physiological substrate of the Ub system (MATalpha2 transcriptional repressor) (1990); cloned the first genes encoding Ub precursors (1984-1989); identified the first Ub-conjugating (E2) enzymes with specific biological functions (RAD6, in DNA repair, and CDC34, in the cell cycle (1987-1988)); discovered the first nonproteolytic function of Ub (as a cotranslational chaperone); identified as well as cloned the first specific E3 Ub ligase, termed UBR1 (1990); and also cloned the first deubiquitylating enzymes, termed UBP1-UBP3. The cloning, in 1990, of the first specific E3 Ub ligase opened up a particularly vast field, as later studies showed that the human genome encodes at least 800 E3 Ub ligases. This multitude of E3s, which recognize distinct degrons in their protein targets, underlies the immense functional reach of the Ub system. In addition, we discovered the first specific poly-Ub chains (in 1989), identified their topology (isopeptide bonds between Gly76 and Lys48 of adjacent Ub moieties), and demonstrated the necessity of these substrate-linked poly-Ub chains for the proteasome-mediated proteolysis. We also discovered that the Ub system is capable of subunit selectivity, i.e., it can destroy a specific subunit of oligomeric protein, leaving the rest of the protein intact and thereby making possible protein remodeling. Subunit selectivity of degradation underlies essential aspects of the cell cycle (selective replacement of cyclin subunits in cell-cycle kinases), the activation of transcription factors such as, e.g., NF-kappaB, and a multitude of other processes. Together, the insights described in items 2 and 3 comprised the discovery of degrons and biological fundamentals of the Ub system.
The Hershko laboratory produced the first of these major advances, in 1978-1983 (item 1), and my laboratory produced the other two, in 1984-1990 (items 2 and 3).
These complementary insights by the two laboratories led to enormous expansion of the Ub field in the 1990s and afterward. This field is now one of the largest arenas in biomedical science, the point of convergence of many disparate disciplines.
The discoveries during the 1980s (items 1-3) yielded the modern paradigm of the central importance of regulated proteolysis for the control of the levels of specific proteins in vivo, as distinguished from their control by transcription and protein synthesis. In sum, these advances revealed that the control through regulated protein degradation rivals, and often surpasses in significance the classical regulation through transcription and translation. This radically changed understanding of the design of biological circuits has major ramifications for medicine, given the vast functional range of the Ub system and the multitude of ways in which Ub-dependent processes can malfunction in disease or in the course of aging, from cancer and neurodegenerative syndromes to perturbations of immunity and many other illnesses, including birth defects.
Papers about early history of the ubiquitin field:
Hershko, A., Ciechanover, A. and Varshavsky, A. (2000) The ubiquitin system. Nature
Medicine 6, 1073-1081.
Varshavsky, A. (2006) The early history of the ubiquitin field. Protein Science 15, 647-654.
Varshavsky, A. (2008) Discovery of cellular regulation by protein degradation. J. Biol. Chem.
283, 34469-34489.
Varshavsky, A. (2014) Discovery of the biology of the ubiquitin system. J. Am. Med.
Association (JAMA) 311, 1969-1970.
Lab's Current and Earlier Studies
In 2007 and also in 2023, we proposed that DNA deletions and (less frequent) insertions that are confined to cancer cells can be used as targets for selective therapy of cancers. Downloadable PDFs of these 2007 and 2023 papers are cited above, after "Research Themes".
My laboratory continues to study the Ub system, with an emphasis on N-degron pathways of protein degradation (they were previously called "N-end rule pathways"), which we analyze in the mouse, in the yeast Saccharomyces cerevisiae, and in bacteria. Although bacteria lack the bona fide Ub system, at least one (Ub-independent) N-degron pathway, termed the Leu/N‑degron pathway (Fig. 1D), is present in them as well.
We also study molecular cause(s) of sleep, i.e., the problem of why we sleep, as distinguished from how we sleep. The understanding of sleep regulation (how we sleep) is already extensive, while the understanding of exactly why we sleep is still to be attained.
In 2012 (Varshavsky, A. (2012) Protein Science, 21:1634-1661), and also later, in 2019 (Varshavsky, A. (2019) PNAS 116, 10773–10782), I proposed that if there's a specific molecular cause of sleep (as distinguished from multiple comparably significant causes), it might stem from production, during wakefulness, of numerous intracellular and extracellular protein fragments that can be transiently beneficial but can also perturb, through their cumulative and mechanistically diverse effects, the functioning of the brain and other organs. A part of the fragment generation (FG) hypothesis is the suggestion that some natural protein fragments are removed too slowly during wakefulness, and that the resulting accumulation of fragments gradually impairs cognition and other processes. In sum, the FG hypothesis posits that sleep evolved, at least in part, to counteract overproduction (owing to insufficiently fast elimination) of hundreds of different protein fragments that may accumulate, at low fractional levels, in the brain and other sleep‑relevant organs (e.g., the skeletal muscle) by the time sleep begins. Our studies in this arena focus on verification of the FG hypothesis.
Early Years of the Laboratory, Before 1984
Our studies in 1978-1981 produced two sets of advances outside the ubiquitin field. One of them was the discovery, in 1978-1979, of the first nucleosome-depleted, nuclease-hypersensitive regions in chromosomes (Varshavsky et al. (1978) SV40 minichromosome: preferential exposure of the origin of replication. Nucl. Acids Res. 5, 3469-3478. Varshavsky et al. (1979) A 400 bp stretch of SV40 viral DNA that includes the origin of replication is exposed in minichromosomes. Cell 16, 453-466). Such regions are characteristic, in particular, of transcriptional promoters, recombinational hotspots, and the origins of DNA replication. This arena continues to be an active field of chromosome studies.
The other advance outside the ubiquitin field was the discovery and dissection, in 1980-1981, of the first pathway of chromosome cohesion/segregation (Sundin & Varshavsky (1980) Terminal stages of SV40 DNA replication proceed via multiply intertwined catenated dimers. Cell 21, 103-114. Sundin & Varshavsky (1981) Arrest of segregation leads to accumulation of highly intertwined catenated dimers: dissection of the final stages of SV40 DNA replication. Cell 25, 659-669). This chromosome cohesion/segregation pathway involves the formation, during replication, of multiply intertwined (multicatenated) sister chromatids, and their subsequent regulated separation, through stepwise decatenation, by topo-II isomerases.
New Methods Introduced by the Laboratory
In addition to ubiquitin/N-degron studies and other contributions described above, the lab invented, over the years, a variety biochemical and genetic methods. These techniques included:
2-D mapping of DNA replication intermediates [1,2];
2-D hybridization mapping of nucleosomes [3];
Ub fusion technique [4,5];
Chromatin immunoprecipitation (ChIP) assay [6];
Hypersensitivity to heavy water as a generally applicable conditional phenotype [7];
Heat-activated degron [8];
Split-Ub method for detecting protein interactions [9];
Ub translocation assay [10];
Ub reference technique [11];
Ub sandwich technique [12];
Subunit decoy method [13];
Promoter reference technique (PRT) [14])
Most of these methods continue to be in use, and some of the cited techniques remain uniquely important in their areas of application.
References to the Section About New Methods:
- Sundin & Varshavsky (1980) Terminal stages of SV40 DNA replication proceed via multiply intertwined catenated dimers. Cell 21, 103-114.
- Sundin & Varshavsky (1981) Arrest of segregation leads to accumulation of highly intertwined catenated dimers: dissection of SV40 DNA replication. Cell 25, 659-669).
- Levinger et al. (1981) Two-dimensional hybridization mapping of nucleosomes. comparison of DNA and protein patterns. J. Mol. Biol. 146, 287-304.
- Bachmair et al. (1986) In vivo half-life of a protein is a function of its amino-terminal residue. Science 234, 179-186.
- Varshavsky, A. (2005) Ubiquitin fusion technique and related methods. Meth. Enzymol. 399, 777-799.
- Solomon et al. (1988) Mapping protein-DNA interactions in vivo with formaldehyde. Cell 53, 937-947.
- Bartel & Varshavsky (1988) Hypersensitivity to heavy water: a new conditional phenotype. Cell 52, 935-941.
- Dohmen et al. (1994) Heat-inducible degron: a method for constructing temperature-sensitive mutants. Science 263, 1273-1276.
- Johnsson & Varshavsky (1994) Split ubiquitin as a sensor of protein interactions in vivo. PNAS 91, 10340-10344.
- Johnsson & Varshavsky (1994) Ubiquitin-assisted dissection of protein transport across cell membranes. EMBO J. 13, 2686-2698.
- Lévy et al. (1996) Using ubiquitin to follow the metabolic fate of a protein. PNAS 93, 4907-4912.
- Turner & Varshavsky (2000) Detecting and measuring cotranslational protein degradation in vivo. Science 289, 2117-2120.
- Shemorry et al. (2013) Control of protein quality and stoichiometries by N-terminal acetylation and the N-end rule pathway. Molecular Cell 50, 540-551.
- Oh et al. (2017) A reference-based protein degradation assay without global translation inhibitors. J. Biol. Chem. 292, 21457-21465.
1984-1990 Papers by the Laboratory That Co-Founded the Ubiquitin Field
Özkaynak, E., Finley, D. and Varshavsky, A. (1984) The yeast ubiquitin gene: head-to-tail repeats encoding a polyubiquitin precursor protein. Nature 312, 663-666.
Finley, D., Ciechanover, A. and Varshavsky, A. (1984) Thermolability of ubiquitin-activating enzyme from the mammalian cell cycle mutant ts85. Cell 37, 43-55.
Ciechanover, A., Finley, D. and Varshavsky, A. (1984) Ubiquitin dependence of selective protein degradation demonstrated in the mammalian cell cycle mutant ts85. Cell 37, 57-66.
Bachmair, A., Finley, D. and Varshavsky, A. (1986) In vivo half-life of a protein is a function of its N‑terminal residue. Science 234, 179-186.
Finley, D., Özkaynak, E. and Varshavsky, A. (1987) The yeast polyubiquitin gene is essential for resistance to high temperatures and other stresses. Cell 48, 1035-1046.
Jentsch, S., McGrath, J. P. and Varshavsky, A. (1987) The yeast DNA repair gene RAD6 encodes a ubiquitin-conjugating enzyme. Nature 329, 131-134.
Goebl, M. G., Jentsch, S., McGrath, J. P., Varshavsky, A. and Byers, B. (1988) The yeast cell cycle gene CDC34 encodes a ubiquitin conjugating enzyme. Science 241, 1331-1335.
Bachmair, A. and Varshavsky, A. (1989) The degradation signal in a short-lived protein. Cell 56, 1019‑1032.
Chau, V., Tobias, J. W., Bachmair, A., Mariott, D., Ecker, D., Gonda, D. K., and Varshavsky, A. (1989) A multiubiquitin chain is confined to a specific lysine in a targeted short‑lived protein. Science 243, 1576-1583.
Finley, D., Bartel, B. and Varshavsky, A. (1989) The tails of ubiquitin precursors are ribosomal proteins whose fusion to ubiquitin facilitates ribosome biogenesis. Nature 338, 394‑401.
Gonda, D. K., Bachmair, A., Wünning, I., Tobias, J. W. and Varshavsky, A. (1989) Universality and structure of the N-end rule. J. Biol. Chem. 264, 16700-16712.
Hochstrasser, M. and Varshavsky, A. (1990) In vivo degradation of a transcriptional regulator: the yeast alpha-2 repressor. Cell 61, 697-708.
Johnson, E. S., Gonda, D. K. and Varshavsky, A. (1990) Cis-trans recognition and subunit specific degradation of short-lived proteins. Nature 346, 287-291.
Bartel, B., Wünning, I. and Varshavsky, A. (1990) The recognition component of the N-end rule pathway. EMBO J. 9, 3179-3189.
Selected Papers, 1991-Present
Hochstrasser, M., Ellison, M. J., Chau, V. and Varshavsky, A. (1991) The short-lived MATalpha2 transcriptional regulator is ubiquitinated in vivo. PNAS 88, 4606‑4610.
Baker, R. T. and Varshavsky, A. (1991) Inhibition of the N-end rule pathway in living cells. PNAS 88, 1090-1094.
Varshavsky, A. (1991) Naming a targeting signal. Cell 64, 13-15.
Tobias, J. W. and Varshavsky, A. (1991) Cloning and functional analysis of the ubiquitin- specific protease gene UBP1 of S. cerevisiae. J. Biol. Chem. 266, 12021-12028.
Dohmen, R. J., Madura, K., Bartel, B. and Varshavsky, A. (1991) The N-end rule is mediated by the Ubc2 (Rad6) ubiquitin-conjugating enzyme. PNAS 88, 7351‑7355.
McGrath, J. P., Jentsch, S. and Varshavsky, A. (1991) UBA1: an essential yeast gene encoding ubiquitin-activating enzyme. EMBO J. 10, 227-237.
Tobias, J. W., Shrader, T. E., Rocap, G. and Varshavsky, A. (1991) The N-end rule in bacteria. Science 254, 1374-1377.
Johnson, E. S., Bartel, B., Seufert, W. and Varshavsky, A. (1992) Ubiquitin as a degradation signal. EMBO J. 11, 497-505.
Baker, R. T., Tobias, J. W. and Varshavsky, A. (1992) Ubiquitin-specific proteases of S. cerevisiae: cloning of UBP2 and UBP3, and functional analysis of the UBP gene family. J. Biol. Chem. 267, 23363-23375.
Varshavsky, A. (1992) The N-end rule. Cell 69, 725-735.
Shrader, T. E., Tobias, J. W. and Varshavsky, A. (1993) The N-end rule in Escherichia coli: cloning and analysis of the leucyl, phenylalanyl-tRNA-protein transferase gene aat. J. Bact. 175, 4364‑4374.
Madura, K., Dohmen, R. J. and Varshavsky, A. (1993) N-recognin/Ubc2 interactions in the N‑end rule pathway. J. Biol. Chem. 268, 12046‑12054.
Ota, I. M. and Varshavsky, A. (1993) A yeast protein similar to bacterial two-component regulators. Science 262, 566-569.
Dohmen, R. J., Wu, P. P. and Varshavsky, A. (1994) Heat-inducible degron: a method for constructing temperature-sensitive mutants. Science 263, 1273-1276.
Johnsson, N. and Varshavsky, A. (1994) Ubiquitin-assisted dissection of protein transport across membranes. EMBO J. 13, 2686-2698.
Johnsson, N. and Varshavsky, A. (1994) Split ubiquitin as a sensor of protein interactions in vivo. PNAS 91, 10340‑10344.
Varshavsky, A. (1995) Codominance and toxins: a path to drugs of nearly unlimited selectivity. PNAS 92, 3663-3667.
Johnston, J. A., Johnson, E. S., Waller, P. and Varshavsky, A. (1995) Methotrexate inhibits proteolysis of dihydrofolate reductase by the N-end rule pathway. J. Biol. Chem. 270, 8172‑8178.
Baker, R. T. and Varshavsky, A. (1995) N-terminal amidase: a new enzyme and component of a targeting complex in the N-end rule pathway. J. Biol. Chem. 270, 12065‑12074.
Varshavsky, A. The world of ubiquitin. (1995) Engineering & Science 58, 26‑36.
Johnson, E. S., Ma, P. C. M., Ota, I. M. and Varshavsky, A. (1995) A proteolytic pathway that recognizes ubiquitin as a degradation signal. J. Biol. Chem. 270, 17442‑17456.
Ghislain, M., Dohmen, R. J., Lévy, F., and Varshavsky, A. (1996) Cdc48p interacts with Ufd3p, a WD-repeat protein required for ubiquitin-dependent proteolysis in S. cerevisiae. EMBO J. 15, 4884-4899.
Varshavsky, A. (1996) The N-end rule: functions, mysteries, uses. PNAS 93, 12142-12149.
Grigoryev, S., Stewart, A. E., Kwon, Y. T., Arfin, S. M., Bradshaw, R. A., Jenkins, N., Copeland, N. G. and Varshavsky, A. (1996) A mouse amidase specific for N-terminal asparagine: the gene, the enzyme, and their function in the N-end rule pathway. J. Biol. Chem. 271, 28521-28532.
Varshavsky, A. (1997) The ubiquitin system. Trends Biochem. Sci. 22, 383-387.
Byrd, C. Turner, G. and Varshavsky, A. (1998) The N-end rule pathway controls the import of peptides through degradation of a transcriptional repressor. EMBO J. 17, 269-277.
Kwon, Y. T., Reiss, Y., Fried, V. A., Hershko, A., Yoon, J. K., Gonda, D. K., Sangan, P., Copeland, N. G., Jenkins, N. A. and Varshavsky, A. (1998) The mouse and human genes encoding the recognition component of the N-end rule pathway.PNAS 95, 7898-7903.
Varshavsky, A. (1998) Codominant interference, antieffectors, and multitarget drugs. PNAS 95, 2094-2099.
Kwon, Y. T., Kashina, A. S. and Varshavsky, A. (1999) Alternative splicing results in differential expression, activity and localization of the two forms of arginyl-tRNA-protein transferase, a component of the N-end rule pathway. Mol. Cell. Biol. 19, 182-193.
Kwon, Y. T. Lévy, F. and Varshavsky, A. (1999) Bivalent inhibitor of the N-end rule pathway. J. Biol. Chem. 274, 18135-18139.
Xie, Y. and Varshavsky, A. (1999) The N-end rule pathway is required for import of histidine in yeast lacking the kinesin-like protein Cin8p. Curr. Genet. 36, 113-123.
Xie, Y. and Varshavsky, A. (1999) The E2-E3 interactions in the N-end rule pathway: the RING‑H2 of E3 is required for the synthesis of multiubiquitin chain. EMBO J. 18, 6832‑6844.
Suzuki, T. and Varshavsky, A. (1999) Degradation signals in the lysine-asparagine sequence space. EMBO J. 18, 6017-6026.
Davydov, I. V. and Varshavsky, A. (2000) RGS4 is arginylated and degraded by the N-end rule pathway in vitro. J. Biol. Chem. 275, 22931-22941.
Hershko, A., Ciechanover, A. and Varshavsky, A. (2000) The ubiquitin system. Nature Medicine 6, 1073-1081.
Turner, G. C. and Varshavsky, A. (2000) Detecting and measuring cotranslational protein degradation in vivo. Science 289, 2117-2120.
Xie, Y. and Varshavsky, A. (2000) Physical association of ubiquitin ligases and the 26S proteasome. PNAS 97, 2497-2502.
Kwon, Y. T., Balogh, S. A., Davydov, I. V., Kashina, A. S., Yoon, J. K., Xie, Y., Gaur, A., Hyde, L., Denenberg, V. H. and Varshavsky, A. (2000) Altered activity, social behavior, and spatial memory in mice lacking the NTAN1p amidase and the asparagine branch of the N-end rule pathway. Mol. Cell. Biol. 20, 4135-4148.
Turner, G. C., Du, F. and Varshavsky, A. (2000) Peptides accelerate their uptake by activating a ubiquitin-dependent proteolytic pathway. Nature 405, 579-583.
Hershko, A., Ciechanover, A. and Varshavsky, A. (2000) The ubiquitin system. Nature Medicine 6, 1073-1081.
Rao, H., Uhlmann, F., Nasmyth, K. and Varshavsky, A. (2001) Degradation of a cohesin subunit by the N‑end rule pathway is essential for chromosome stability. Nature 410, 955‑960.
Xie, Y. and Varshavsky, A. (2001) RPN4 is a ligand, substrate, and transcriptional regulator of the 26S proteasome: a negative feedback circuit. PNAS 98, 3056‑3061.
Kwon, Y. T., Xia, Z.-X., Davydov, I. V., Lecker, S. H. and Varshavsky, A. (2001) Construction and analysis of mouse strains lacking the ubiquitin ligase UBR1 of the N-end rule pathway. Mol. Cell. Biol. 21, 8007-8021.
Kwon, Y. T., Kashina, A. S., Davydov, I. V., Hu, R.-G., An, J. Y., Seo, J. W., Du, F. And Varshavsky, A. (2002) An essential role of N-terminal arginylation in cardiovascular development. Science 297, 96-99.
Du, F., Navarro-Garcia, F. Xia, Z., Tasaki, T. and Varshavsky, A. (2002) Pairs of dipeptides activate the binding of substrate by ubiquitin ligase through dissociation of its autoinhibitory domain. PNAS 99, 14110-14115.
Sheng, J., Kumagai, A., Dunphy, W. and Varshavsky, A. (2002) Dissection of c-MOS degron. EMBO J. 21, 6061-6071.
Xie, Y. and Varshavsky, A. (2002) The UFD4 ubiquitin ligase lacking the proteasome‑binding region catalyzes ubiquitylation but is impaired in proteolysis. Nature Cell Biol. 4, 1003-1007.
Varshavsky, A. (2003) The N-end rule and regulation of apoptosis. Nature Cell Biol. 5, 373‑376.
Kwon, Y. T., Xia, Z., An, J. Y., Tasaki, T., Davydov, I. V., Seo, J. W., Sheng, J., Xie, Y. and Varshavsky, A. (2003) Female lethality and apoptosis of spermatocytes in mice lacking the UBR2 ubiquitin ligase of the N-end rule pathway. Mol. Cell. Biol. 23, 8255-8271.
Varshavsky, A. (2004) Spalog and sequelog: neutral terms for spatial and sequence similarity. Curr. Biol. 14, R181-R183.
Varshavsky, A. (2005) Regulated protein degradation. Trends Biochem. Sci. 6, 283-286.
Tasaki, T., Mulder, L. C. F., Iwamatsu, A., Lee, M. J., Davydov, I. V., Varshavsky, A., Muesing, M. and Kwon, Y. T. (2005) A family of mammalian E3 ubiquitin ligases that contain the UBR motif and recognize N‑degrons. Mol. Cell. Biol. 25, 7120-7136.
Hu, R.G., Sheng, J., Qi, X., Xu, Z., Takahashi, T. T. and Varshavsky, A. (2005) The N-end rule pathway as a nitric oxide sensor controlling the levels of multiple regulators. Nature 473, 981-986.
Zenker, M, Mayerle, J., Lerch, M. M., (et al.), Kwon, Y. T., Varshavsky, A. and Reis, A. (2005) Deficiency of UBR1, a ubiquitin ligase of the N‑end rule pathway, causes pancreatic dysfunction, malformations and mental retardation (Johanson-Blizzard syndrome). Nature Genet. 37, 1345-1350.
Graciet, E. Hu, R. G., Piatkov, K., Rhee, J. H., Schwarz, E. M. and Varshavsky, A. (2006) Aminoacyl-transferases and the N-end rule pathway of prokaryotic/eukaryotic specificity in a human pathogen. PNAS 103, 3078‑3083.
An, J. Y., Seo, J. W., Tasaki, T., Lee, M. J., Varshavsky, A., Kwon, Y. T. (2006) Impaired neurogenesis and cardiovascular development in mice lacking the E3 ubiquitin ligases UBR1 and UBR2 of the N-end rule pathway. PNAS 103, 6212‑6217.
Varshavsky, A. (2006) The early history of the ubiquitin field. Protein Science 15, 647-654.
Hu, R.-G., Brower, C. S., Wang, H., Davydov, I. V, Sheng, J., Zhou, J., Kwon, Y. T. and Varshavsky, A. (2006) Arginyl-transferase, its specificity, putative substrates, bidirectional promoter, and splicing-derived isoforms. J. Biol. Chem. 281, 32559-32573.
Tasaki, T., Sohr, R., Hellweg, R., Hortnagl, H., Varshavsky, A. and Kwon, Y. T. (2007) Biochemical and Genetic Studies of UBR3, a Ubiquitin Ligase with a Function in Olfactory and Other Sensory Systems. J. Biol. Chem. 282, 18510-18520.
Varshavsky, A. (2007) Targeting the absence: homozygous DNA deletions as immutable signposts for cancer therapy. PNAS 104, 14935-14940.
Hu, R.-G., Wang, H., Xia, Z. and Varshavsky, A. (2008) The N-end rule pathway is a sensor of heme. PNAS 105, 76-81.
Varshavsky, A. (2008) The World as Physics, Mathematics and Nothing Else. In: Physical Biology: From Atoms to Medicine (A. Zewail, ed.), pp.7-22, Imp. College Press, U.K.
Xia, Z., Webster, A., Du, F., Piatkov, K., Ghislain, M. and Varshavsky, A. (2008) Substrate binding sites of UBR1, the ubiquitin ligase of the N-end rule pathway. J. Biol. Chem. 283, 24011-24028.
Hwang, C. S. and Varshavsky, A. (2008) Regulation of peptide import through phosphorylation of Ubr1, the ubiquitin ligase of the N-end rule pathway. PNAS 105, 19188-19193.
Varshavsky, A. (2008) Discovery of cellular regulation by protein degradation. (Reflections article) J. Biol. Chem. 283, 34469-34489.
Hwang, C. S., Shemorry, A. and Varshavsky, A. (2009) Two proteolytic pathways regulate DNA repair by cotargeting the Mgt1 alkylguanine transferase. PNAS 106, 2142-2147.
Brower, C. S. and Varshavsky, A. (2009) Ablation of arginylation in the N-end rule pathway: loss of fat, increased metabolic rate, damaged spermatogenesis, and neurological perturbations. PLoS ONE 4, e7757.
Wang, H., Piatkov, K. I., Brower, C. S. and Varshavsky, A. (2009) Glutamine-specific N-terminal amidase, a component of the N-end rule pathway. Molecular Cell 34, 686-695.
Hwang, C. S., Shemorry, A. and Varshavsky, A. (2010) N-terminal acetylation of cellular proteins creates specific degradation signals. Science 327, 973-977.
Hwang, C. S., Shemorry, A. and Varshavsky, A. (2010) The N-end rule pathway is mediated by a complex of the RING-type Ubr1 and HECT-type Ufd4 ubiquitin ligases. Nature Cell Biol. 12, 1177-1185.
Varshavsky, A. (2011) The N-end rule pathway and regulation by proteolysis. Protein Science 20, 1298-1345.
Varshavsky, A. (2012) The ubiquitin system, an immense realm. Annu. Rev. Biochem. 81, 167-176.
Piatkov, K. I., Brower, C. S. and Varshavsky, A. (2012) The N-end rule pathway counteracts cell death by destroying proapoptotic protein fragments. PNAS 109, E1839-E1847.
Varshavsky, A. (2012) Augmented generation of protein fragments during wakefulness as the molecular cause of sleep: a hypothesis. Protein Science 21, 1634-1661.
Piatkov, K. I., Colnaghi, L., Bekes, M, Varshavsky, A. and Huang, T. (2012) The auto-generated fragment of the Usp1 deubiquitylase is a physiological substrate of the N-end rule pathway. Molecular Cell 48, 926-933.
Brower, C. S., Piatkov, K. I. and Varshavsky, A. (2013) Neurodegeneration-associated protein fragments as short-lived substrates of the N-end rule pathway. Molecular Cell 50, 161-171.
Shemorry, A., Hwang, C. S. and Varshavsky, A. (2013) Control of protein quality and stoichiometries by N-terminal acetylation and the N-end rule pathway. Molecular Cell 50, 540-551.
Kim, H.-K., Kim, R. R. Oh, J.-H, Cho, H., Varshavsky, A. and Hwang, C. S. (2014) The N‑terminal methionine of cellular proteins as a degradation signal. Cell 156, 158-169.
Piatkov, K.I., Oh, J. H., Liu, Y. and Varshavsky, A. (2014) Calpain-generated natural protein fragments as short-lived substrates of the N-end rule pathway. PNAS 111, E817-E826.
Varshavsky, A. (2014) Discovery of the biology of the ubiquitin system. J. Am. Med. Association (JAMA) 311, 1969-1970.
Brower, C. S., Rosen, C. E. Jones, R. H. Wadas, B. C., Piatkov, K. I. and Varshavsky, A. (2014) Liat1, an arginyltransferase-binding protein whose evolution among primates involved changes in the numbers of its 10-residue repeats. PNAS 111, E4936–E4945.
Park, S.-E., Kim, J.-M., Seok, O.H., Cho, H., Wadas, B., Kim, S. Y., Varshavsky, A. and Hwang, C. S. (2015) Control of mammalian G protein signaling by N-terminal acetylation and the N-end rule pathway. Science 347, 1249-1252.
Piatkov, K. I., Vu, T. T. M., Hwang, C. S. and Varshavsky, A. (2015) Formyl-methionine as a degradation signal at the N-termini of bacterial proteins. Microbial Cell 2, 376-393.
Wadas, B., Borjigin, J., Huang, Z. Oh, J. H., Hwang, C. S. and Varshavsky, A. (2016) Degradation of serotonin N-acetyltransferase, a circadian regulator, by the N-end rule pathway. J. Biol Chem. 291, 17178-17196.
Wadas, B., Piatkov, K. I., Brower, C. S. and Varshavsky, A. (2016) Analyzing N-terminal arginylation through the use of peptide arrays and degradation assays. J. Biol. Chem. 291, 20976-20992.
Chen, S. J., Wu, X., Wadas, B., Oh, J. H. and Varshavsky, A. (2017) An N-end rule pathway that recognizes proline and destroys gluconeogenic enzymes. Science 355, eaal3655.
Oh, J. H., Hyun, J. Y. and Varshavsky, A. (2017) Control of Hsp90 chaperone and its clients by N-terminal acetylation and the N-end rule pathway. PNAS 114, E4370-E379.
Dougan, D. A. and Varshavsky, A. (2018) Understanding the Pro/N-end rule pathway. Nature Chemical Biology 14, 415-416.
Kim, J. M., Seok, O. H., Ju, S., Heo, J. E., Yeom, J., Kim, D. S., Yoo, J. Y., Varshavsky, A., Lee, L. and Hwang., C. S. (2018) Formyl-methionine as an N-degron of a eukaryotic N‑end rule pathway. Science 362, eaat1074.
Varshavsky, A. (2019) N-degron and C-degron pathways of protein degradation. PNAS 116, 358-366.
Varshavsky, A. (2019) On the cause of sleep: protein fragments, the concept of sentinels, and links to epilepsy. PNAS 116, 10773-10782.
Melnykov, A., Chen, S. J. and Varshavsky, A. (2019) Gid10 as an alternative N-recognin of the Pro/N-degron pathway. PNAS 116, 15914–15923.
Chen, S. J., Melnykov, A. and Varshavsky, A. (2020) Evolution of substrates and components of the Pro/N-degron pathway. Biochemistry 59, 541-551.
Oh, J. H., Chen, S. J. and Varshavsky, A. (2017) A reference-based protein degradation assay without global translation inhibitors. J. Biol. Chem. 292, 21457-21465.
Oh, J. H., Hyun, J. Y., Chen, S. J. and Varshavsky, A. (2020) Five enzymes of the Arg/N‑degron pathway form a targeting complex: the concept of superchanneling. PNAS 117, 10778-1788.
Vu, T. T. M and Varshavsky, A. (2020) The ATF3 transcription factor is a short-lived substrate of the Arg/N-degron pathway. Biochemistry 59, 2796-2812.
Vu, T. T. M., Mitchell, D. C., Gygi, S. P. and Varshavsky, A. (2020) The Arg/N-degron pathway targets transcription factors and regulates specific genes. PNAS 117, 31094-31104.
Chen S. J., Kim, L., Song, H. K. and Varshavsky, A. (2021) Aminopeptidases trim Xaa-Pro proteins, initiating their degradation by the Pro/N-degron pathway. PNAS 118, e2115430118.
Kim, B. H., Kim, M. K., Oha, S. J., Nguyen, K. T., Kima, J. H. Varshavsky, A., Hwang, C. S. and Song, H. K. (2022) Crystal structure of the Ate1 arginyl-tRNA-protein transferase and arginylation of N-degron substrates. Proc. Natl. Acad. Sci. USA 119, e2209597119.
Varshavsky, A., Lewis, K. and Chen, S. J. (2023) Deletions of DNA in cancer and their possible use for therapy. BioEssays 2300051.